Varför har människan och alla andra primater ögonen riktade rakt framåt och ett »chiasma opticum« (synnervskors, OC) som nästan till hälften (45 procent) består av okorsade banor? Den traditionella förklaringen är att sådana synbanor tillsammans med höggradig binokularitet befrämjar djupseende [1]. En översiktsartikel i Brain, Behavior and Evolution [2] för fram en ny »Eye-forelimb-hypothesis« (EF-hypotes) som föreslår att den speciella neurala arkitekturen hos primater utvecklats av ett helt annat skäl. EF-hypotesen postulerar att det har selektivt värde att ha korta nervbanor mellan områden i hjärnan som mottar syninformation om handen och de motorkärnor som styr handens koordination.
Grundläggande för EF-hypotesen är att evolutionära förändringar i OC kommer att påverka längden av dessa nervbanor. Figur B här intill visar att primatens anatomi med en hög andel (okorsade) ipsilaterala retinala projektioner (IRP) medför att all visuell information om till exempel högra handen kommer till »rätt sida«, det vill säga vänster hemisfär, så länge handen befinner sig till höger om medellinjen. Denna anatomi bör ha adaptivt värde hos djur som primater, som ofta använder den främre extremiteten framåt i synfältet, medan en låg andel IRP bör ha adaptivt värde hos djur som mestadels eller alltid använder den främre extremiteten inom det laterala synfältet (se figur A). Även om teoretiska modeller och anatomiska figurer ter sig övertygande bör man fråga sig om det finns något experimentellt stöd. Ett sätt att pröva hypotesen är att jämföra precision och hastighet för, låt oss säga, högra handen när den utför uppgifter i höger respektive vänster synfält. Flera sådana experiment har gjorts (dock inte i syfte att pröva EF-hypotesen). Resultaten talar för det som EF-hypotesen postulerar, nämligen en högre precision och hastighet så länge handen arbetar i det ipsilaterala synfältet [3-5]. Berlucchi et al [3] menar att handens reaktioner på visuella stimuli presenterade i det ipsilaterala synfältet integreras i den kontralaterala hemisfären och att det ger färre synapser att passera för nervsignalerna, vilket resulterar i snabbare motorik än med visuella stimuli som presenteras kontralateralt.
EF-hypotesen evalueras även genom komparativa analyser, bland annat relateras olika djurs grad av IRP till synfältens utformning, ekologisk nisch och motorbeteende [2]. Bland vertebrater utanför däggdjursgruppen har variationerna i IRP tidigare ansetts vara oförklarliga [6]. En rad anatomiska, neurofysiologiska, fylogenetiska, ontogenetiska och ekologiska data läggs fram till stöd för EF-hypotesen.
Primater och kattdjur har en stark koppling mellan ögats och den främre extremitetens motorik. Katter har i likhet med primater en hög andel IRP (30 procent). Krokodiler, flertalet fåglar och fiskar saknar IRP [6]. Sistnämnda djur har av anatomiska/funktionella skäl knappast någon användning för främre extremiteten i frontalt läge. Den afrikanska klogrodan Xenopus laevis har enbart korsade projektioner fram till metamorfosen, varvid den utvecklar binokulärt seende och främre extremiteter med klor. X laevis använder klorna när den fångar bytesdjur framför sig. Delfiner saknar IRP, vilket är i överensstämmelse med hypotesen eftersom delfinens främre extremitet (pektoralfenan) används enbart i lateralt läge. Bland pungdjuren (Marsupialia) har tre trädklättrande arter en hög andel IRP medan vombater (Vombatidae), en marklevande växtätare som rimligen har mindre behov av visuell styrning av framfoten, har minst IRP. Övriga pungdjur faller mellan dessa extremer [2].
Detta är till sin karaktär grundforskning som i första hand kan bidra till ökad förståelse kring hjärnans evolution, och forskning om öga och hand är ett stort och aktivt forskningsområde. Kan EF-hypotesen tänkas få någon klinisk betydelse? Datorn är vårt vanligaste arbetsredskap. Vid arbete med mus krävs en god öga–handkoordination, och många har besvär som kan kopplas till sådant arbete. Ökad förståelse av hur samverkan mellan synsystem och rörelseorgan fungerar kan få stor betydelse för individ och samhälle, inte minst om det leder till bättre förebyggande insatser och behandlingar. Kanske kan EF-hypotesen även bidra till insikter om patofysiologin vad gäller rörelseorganen i nacke/skuldra.
EF-hypotesen ger nya perspektiv på människans evolution. God öga–handkontroll är typiska drag hos primater [7]. Fossilfynd talar för att de första egentliga primaterna uppträdde för ungefär 55 miljoner år sedan. Redan då tycks handen ha varit specialiserad för att gripa med. Urprimater kan ha utvecklat denna speciella greppförmåga för att kunna äta blommor, nektar och löv i trädens distala grenar [3, 4]. Bloch och Boyer gör gällande att handens gripförmåga utvecklades före primaternas visuella specialisering [8]. EF-hypotesen indikerar däremot att primaternas synsystem utvecklades parallellt med handens specialisering genom en gemensam evolutionär mekanism, nämligen att en ökning av IRP hade selektivt värde hos djur som regelbundet använde främre extremiteten i det frontala synfältet.
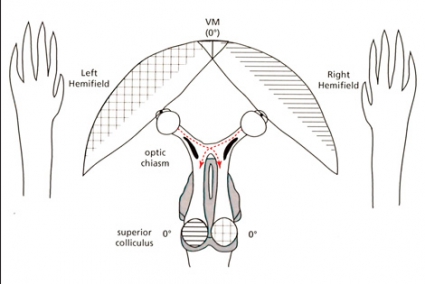
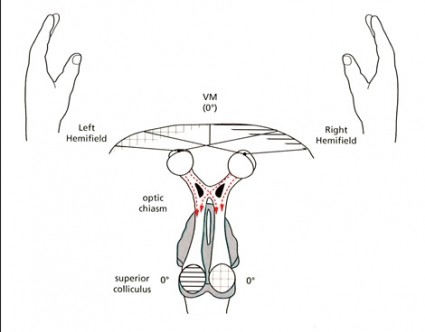
Synbanor hos 2 vertebrater. Hos ryggradsdjur är extremiteternas motorkärnor och somatosensoriska områden belägna i den kontralaterala hemisfären. Figuren visar synbanorna till colliculus superior (CS) hos två principiellt olika typer av djur, ett ryggradsdjur med lateralt placerade främre extremiteter (till exempel en fisk, krokodil, fågel eller mus), A, samt en primat, B. CS är en komponent av mesencefalon (mitthjärnan), som integrerar visuell, somatosensorisk och akustisk information, men principen om nervbanornas längd blir densamma för primär synbark. Figurerna visas med tillstånd från Brain, Behavior and Evolution (Karger). Figurerna är modifierade från Todd Preuss [9] med vänligt tillstånd och bygger i sin tur på en figur framställd av Jack Pettigrew et al [10].